两个或多个原子以太旋涡通过涡管相吸作用及范德华力,相互结合在一起,原子以太旋涡外围的以太形成次生以太涡流,包裹原子以太旋涡,让多原子以太旋涡之间的结构保持稳定时空结构,即为分子以太旋涡。涡管相吸作用分同旋异极吸附作用与异旋同极吸附作用,这是共价键的真正内涵,而非西方科学理论描绘下的“共用电子对”这一联结方式,这已经在《万物意志篇》的“耦合结构形态”章节作充分说明。
由于分子耦合结构需要全新的研究才能考查分子以太旋涡的实际形态,这里为了方便描绘,引用的分子结构形态,都是经典化学分子结构及分子式,只是读者理解这种结构时,要自行代入分子以太旋涡耦合结构及分子振动波形态。
一个细胞内部空间,除了离子之外,就是无数的分子以太旋涡填充其空间。应一个分子以太旋涡包含的原子以太旋涡的数量不同,及原子以太旋涡相互结合后空间形态的差异,细胞内部空间,可以粗略分几种分子结构形态,即双原子分子,极性分子,多原子分子,多原子长链分子,长链分子团。这些不同结构形态的分子以太旋涡,其周边空间,都存在波流一体运动形态,这里一一作简单介绍。
1、双原子分子
双原子结合下的分子以太旋涡,必是异旋同极吸附结构,即分子以太旋涡,是由一个顺旋原子以太旋涡与一个逆旋原子以太旋涡通过两边同极涡管相互吸附在一起。
双原子结构下的分子以太旋涡,其空间存在两个原子以太旋涡的振动力场与流场。这两个振动力场,应双方波动的相位不同,而相互削弱对方的振动力场,表现为相互干扰,进而导致形成的流场,也是相互中和或处于最弱能量状态。如此导致周边空间以太缺少稳定单向的牵引作用而形成紊流、湍流运动,包裹着双原子以太旋涡。
一般双原子气体分子,特别是同元素原子的双原子气体,是有最弱的次生以太旋涡能量状态,如O2、H2、N2,等等。

2、极性分子
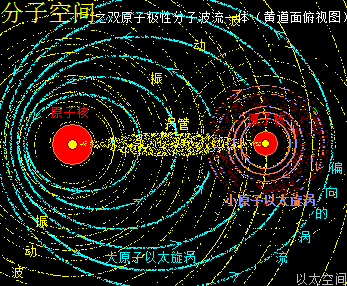
若顺逆互绕的两原子以太旋涡的旋涡角动量不同,振动力场、流场强度相差大,会导致整个分子以太旋涡有很强的偏向性,即极性,在宏观上表现出酸、碱等属性,或氧化性、腐蚀性,等等作用形态,如HCl,NaCl,KCl,OH-,等等。这些分子或离子之中,阳离子的时空尺度与阴离子的时空尺度、力场强度相差较大,会出现类大电子结构下的原子以太旋涡运动形态。
设阳离子的振动力场与流场强度均小于阴离子。如此阴阳离子相互耦合结构下的分子以太旋涡里,其阳离子,就会象阴离子的远核外大正电子以太旋涡,让整个分子以太旋涡的运动形态,有很大的偏向性——这种偏向性要小于游离态的阴离子的偏向性,但要大于同元素原子结构下的双原子分子以太旋涡的偏向性。整个分子以太旋涡,阴离子的振动力场与流场占主导地位,并向外界表达运动特征。阳离子的振动力场与流场,则被包含在阴离子的振动力场与流场之中,处于从属作用地位。

反之,若阳离子的振动力场与流场强度大于阴离子的的振动力场与流场强度,则阴离子会象阳离子的远核外大负电子以太旋涡,也让整个分子以太旋涡的运动形态,有很大的偏向性。整个分子以太旋涡,阳离子的振动力场与流场占主导地位,并向外界表达运动特征。阴离子的振动力场与流场,则被包含在阳离子的振动力场与流场之中,处于从属地位。

这种极性现象,在含有H+与OH-的分子以太旋涡中犹为明显,在于H,即氢原子,是所有元素原子以太旋涡中原子量与时空尺度都最小的原子以太旋涡,非常容易被其它大原子以太旋涡捕获,成为大原子的远核外类大电子。当这种远核外类大电子绕大原子以在旋涡作高速核外运动时,产生最强的以太涡流偏向作用与振动波传递,从而对分子以太旋涡外围空间的其它物质结构产生力的作用,由此导致其它物质产生化学反应或时空结构形态形变,是分子以太旋涡表达出酸性、碱性或其它特殊物质作用的时空结构形态根源所在。特别是在生命体的基础分子结构中,如氨基酸、核苷酸,都是由于含有大量的H+、HO-两种离子而有特殊的物质作用形态,也是中草药对人体产生热性、寒性感观的的内因,会在后面小节穿插说明这两种离子的一般特性。
3、多原子分子
多原子分子,就是构成的原子以太旋涡数量在三个以上,在几十个之内,可以通过简单分子式描绘出的分子以太旋涡。无机分子或离子如H2O、H2SO4、CaCO3、CO2、NO3-,等等,或有机物分子如氨基酸分子,核苷酸分子,羧基、羧酸,烃基分子,等等。
这些分子以太旋涡,也是通过涡管吸附、范德华力相互结合在一起,形成的多旋涡空间嵌套、多旋涡互绕的分子形态。由于这种分子是由可数的不同原子量、振动力场涡与流场强度的原子以太旋涡组合而成,分子中的单个原子以太旋涡对整个分子的影响明显,特别是包含大原子以太旋涡,且原子数量又少的时候,会起主导影响,这是区分多原子分子与长链分子团的一个重要特征。
这种多原子分子团,由一个大原子以太旋涡或少数几个次大原子以太旋涡主导整个分子以太旋涡的运动形态,并通过整体的振动力场与流场对外产生影响作用,如H2SO4,整体能表达出硫元素原子以太旋涡的作用特征,CaCO3,整体能表达出钙元素原子以太旋涡的作用特征,又如氨基酸分子,整体能表达出氮元素原子以太旋涡的作用特征,等等。
一个多原子分子以太旋涡的振动力场,是由这个分子的所有原子以太旋涡的振动力场相互作用后的形态,是一种相互承载、相互干扰、相互叠加后的力场。由这种振动力场牵引而成的流场,也是一种复杂化的强化或削弱后的以太流场形态。同时,应不同元素原子成分与各原子的方位结构差异,在这个分子的某个或某几个方向表达出局部涡流偏向性,即极性。特别是在有机物的分子中,许多分子含有—H、—HO两种基团,这两种基团会成为分子中的类远核外大电子,再叠加上这种分子以太旋涡本身的极性带来的涡流偏向与力场作用,更进一步强化H+、OH-的作用效果。
如OH-,即氢氧根,在有机物分子中被另描绘为分子式—OH,即羟基,并被经典化学理论认为是10个电子的OH-再失去一个核外电子成为9个电子的—OH,才表现出更强的氧化性。其实OH根本就没有失去电子,也不是10个核外电子。两者结构完全一致,都是氧原子以太旋涡嵌套氢原子以太旋涡这一时空结构。只是在有机物分子中,OH以太旋涡叠加上有机物分子以太旋涡的极性而强化了作用效果而已。
这种叠加效果,就如古代骑兵的马刀(OH)劈砍动作,在马匹(有机物分子)的高速奔跑后(极性)的叠加效果,让马刀有更高的速度产生更强力的劈砍作用。而单就骑兵的马刀(OH)的劈砍动作而言,骑不骑马是完全一致的。宏观与微观的物质作用原理是相通的。

4、长链分子
长链分子,是由多个原子或多个多原子分子串连起来的分子以太旋涡形态。这个串连方式,就如许多颗佛珠通过一根绳子穿过中心的小孔连接起来一样。长链分子主体上是同旋异极吸附结构,是一个原子以太旋涡的南极与另一个原子以太旋涡的北极相吸,这个原子以太旋涡的北极,又与还一个原子以太旋涡的南极相吸,如此反复连接,可以构成极长的分子链。
若这个长链分子与其它长链分子通过范德华力相互吸引、交织,让整个长链分子如一张渔网般铺开,形成空间立体分布形态,这其实就是普通固体的一般结构模式。最直观的就是塑料纤维交织在一起形成网状、膜状结构的形态。在细胞空间内部,也存在大量的这种长链分子以太旋涡结构形态,如肽链、DNA之类的长链分子,链上的原子以太旋涡的数量可达几十万甚至更多。
这种长链分子,由于其长链形空间结构的特殊性,有其特定的两种形态的以太旋涡的振动力场与流场:
一是在长链分子内部,由于原子以太旋涡振动,会在纵向上产生波传递与振动力场。
结果导致整个长链分子,如长绳被抖动后产生绳波一般,而在长链分子上,也产生波传递,这种波传递会影响各原子以太旋涡间的位置关系,从而影响长链分子的空间扭曲形态,进而影响整个长链分子以太旋涡的横向振动波发散形态与振动力场形态。
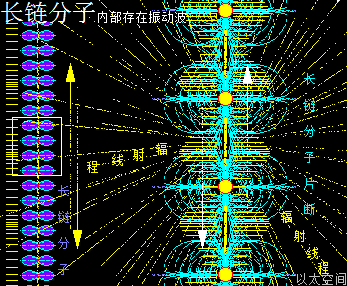
二是在长链分子外部,长链分子里的所有原子以太旋涡或多原子分子以太旋涡的振动力场向四周发散,导致周边空间的以太运动分布形成以分子链为中心线的流场。两者是为波流一体。
这种振动力场,也是一种相互承载、相互干扰、相互叠加后的力场,并叠加有原子以太旋涡、多原子分子以太旋涡的极性,且复杂度更高,让长链分子在局部上具有更广泛的运动形态,可以大规模、批量化实现某种具体的物质作用。
5、长链分子团
长链分子,由于其内部振动波传递与原子以太旋涡的涡流运动,导致原子以太旋涡之间的位置关系不平衡,从而出现扭曲运动。当分子片断持续扭曲相互靠近,通过范德华力相互吸引靠近,最终形成一个毛线团状的缠绕空间结构,就是长链分子团。在细胞空间,存在大量这种长链分子团,如蛋白质分子、染色体,等等。
这种长链分子团,同样由于其团形空间结构的特殊性,也有其特定的两种形态的以太旋涡的振动力场与流场:
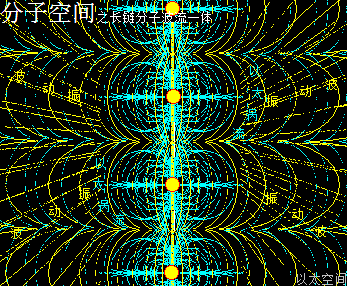
一是在长链分子团内部,由于众多原子以太旋涡振动并在长链上传递,会在团形空间内产生螺旋波传递,即场涡,与涡形振动力场。这个振动力场驱动内部以太流转,两者波流一体。
这内部的振动力场,同时还调整个团形空间里原子以太旋涡的分布形态,最终达成一种分子团空间结构的圆满状态,即团球结构,及分子团内部能量分布的圆满状态,即氲氤状态。这是一个动态的不断的作用过程,并会因外界的干扰而产生对应的形变与重新调整。
二是在长链分子团外部,分子团里的所有原子以太旋涡或多原子分子以太旋涡的振动力场向四周发散,导致周边空间的以太运动分布形成以分子团为中心的流场,两者是为波流一体。
长链分子团的整体作用特征,就是通过内部场涡驱动以太旋涡,犹如一颗行星内部振动驱动周边空间的以太形成以太旋涡。如此一个分子团周边空间,也是存在两极涡管通道结构,与类黄道面的以太旋涡流场。

同时,由于元素原子种类的有限性,与长链分子巨大数量的关系,导致长链分子上存在众多时空结构相近或相同的多原子分子以太旋涡,这些分子以太旋涡之间由于分子团结构而距离过于靠近,当各自涡流的周期性偏向作用而产生振动波后,在相同的方位上会波干涉。干涉波以螺旋涡形自内向外呈辐射形态传递,继而影响外围空间其它分子以太旋涡的分布形态。
这种螺旋涡形辐射形态的干涉波,是细胞形成血管、脊柱、肠道等等圆柱空间结构的重要作用因素,会在后续章节中陆续说明。
整个细胞、器官、人体空间,就是由这些众多离子、双原子分子、多原子分子、长链分子、长链分子团等不同时空尺度的微观以太旋涡相互吸引、堆积而成。各原子、分子以太旋涡,在建构出更高时空尺度的同时,不断展现出原子以太旋涡振动力场与流场的空间运动形态,也与太阳系的物质结构与运动形态相近,这就是全息。全息建构模式,体现在生命的各个时空尺度,也会在后续章节中展现。

由于现代化学及生物学建立的基础理论是在错误的经典原子模型之上的,由此产生的电子云、化学键、电子跃迁发光等概念与原理都是要重新修正、摈弃、批判的,于是当下所有人们在教科书上或学术论文里接受的各类无机化学分子式与有机化学分子式及其对应的分子结构式,乃至复杂的高分子结构式,都须要重新考证与考查其以太旋涡理论下的正确结构模型。这是一个复杂且巨量的人类研究工程,其工作量远超出作者的能力范围之外。因此在以生命运动机制为论述核心的本书各章节里,作者并不尝试纠正已经广为流传的各类无机与有机化学分子式及分子结构式,只是就其主体的结构特征,及未被人们认识的另类运动机制作即生命体各时空尺度下的组织结构的以太旋涡波流一体形态,也即以太旋涡的振动力场与流动力场的统一形态,作一个简单粗略的补充说明。
本书中作者引用所有高分子化学式,如后面的氨基酸分子结构式、核糖核苷酸分子及其它分子结构式等,都不代表作者认可这些化学分子结构式是正确的,在于人类观察能力是不能证明分子结构式的确是这样的。分子式更多的是反映元素原子成分及其比例,这在实验室可以通过宏观的结果产物的比例来证明,而分子结构式则基本是在这成分及比例基础上,通过经典核外电子分布规律、共价键理论、电子云等概念想象出来的,存在大概率甚至完全错误的可能,但为方便说明问题而作一个引用与以太旋涡理论下的补充。更接近客观事实的分子结构式,需待后人的努力与全新观察方法的探寻。读者在理解作者论述时务必要注意这一点,而不是简单相信作者的表述是完全符合客观物质结构形态的。