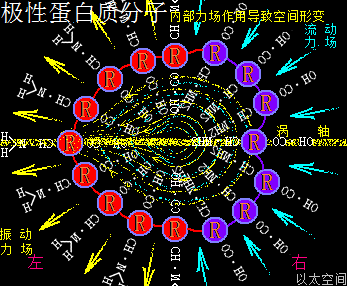
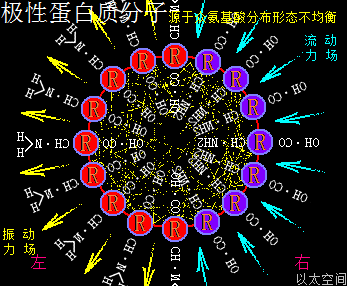
应包含各氨基酸的酸碱性数量不同,及各酸碱性氨基酸在蛋白质颗粒空间的疏密分布不同,导致蛋白质局部空间的振动力场与流动力场分布不均,从而让蛋白质分子次生以太旋涡表现出极性,这与极性分子以太旋涡的原理一样,只是构架空间要广大些,参与的微观以太旋涡要多些。在蛋白质分子空间里,由于氨基酸种类不一,力场不同,导致绝对均衡对称的氨基酸分子分布形态下的蛋白质分子结构是不存在的,于是可知所有蛋白质分子都是极性分子。其以太波流一体状态下的力场形态为:
振动力场、流动力场各自不对称
以一个球形蛋白质分子为例,设分子一半组成是强酸性氨基酸分子,分布在左半边,一半组成是强碱性氨基酸分子,分布在右半边,其振动力场与流动力场整体对外表现为中性,是为中性蛋白质分子。但在局部区域,即接近左半边顶点或右半边顶点的局部空间里,其振动力场与流动力场分别占主导地位。而接近左右半边交接处的局部空间里,其振动力场与流动力场又是相互中和。

如此结构的蛋白质分子,会应外界振动力场与流动力场的作用,而出现快速轴纠正反应。比如这蛋白质分子若出现在一个强振动力场环境空间里,蛋白质分子的振动力场与环境振动力场相互排斥,左半边会被推至背离环境振动源的方向,而右半边会被吸引至靠近环境振动源的方向,表达为轴纠正。反之这蛋白质分子若出现在一个强流动力场环境空间里,蛋白质分子的流动力场与环境流动力场相互排斥,右半边会被推至远离环境振动源的方向,而左半边会被吸引至接近环境振动源的方向,也表达为轴纠正。两种轴纠正都是极性蛋白质分子对环境变化作出的动作反应。即:
极性蛋白质分子会有轴纠正运动
若这个蛋白质分子处于氲氤状态下的以太空间里,由于左半边的振动力场以波的形式产生作用,会在左半边外围空间产生正以太压力,正以太压力会推动蛋白质分子向振动力场的反方向位移,也即右移动。同时,右半边的流动力场以流的形式产生作用,会在右半边外围空间产生负以太压力,负以太压力会牵引此蛋白质分子向流动力场的反方向位移,也即向右移动。两种力场都对此蛋白质分子产生向右的作用力,这蛋白质分子就会向右方向定向位移。自然,由于蛋白分子的振动力场及流动力场同以太空间相互作用的效果是很弱的,这种位移速度也是非常缓慢,但又是存在的。不同极性的蛋白质分子的运动形态也是千差万别的。这是极性蛋白质分子能够自主运动的力场作用根源,而非当下科学界对蛋白质分子的认识只是随机分布在空间的颗粒形态,可以任由环境变化摆布与安排。即:
极性蛋白质分子能够自主定向运动
极性蛋白质分子即能进行轴纠正运动,又能自主定向运动,因此对环境力场变化能随时随地产生空间位置变动与自身状态调整,这就是蛋白质分子的应激反应。
在这球形蛋白质分子内部空间,—CO—OH与—CH—NH2也是对应地处于分布疏密不均的状态,与外部的基键分布刚好相反,导致内部空间的振动力场与流场也是有单一指向性,是从—CH—NH2指向—CO—OH。而—CH—NH2的振动力场有向外扩张的趋势,—CO—O的流动力场有向内收敛的趋势,内部振动波从—CH—NH2指向—CO—OH并形成场涡,再牵引以太形成次生以太旋涡与涡管,涡轴是两力场的中心连接线。于是两个力场分别让—CO—OH聚集的区域向内收敛,又从涡管处向外扩张;让—CH—NH2聚集的区域向外膨胀,又从涡管处向内凹陷,形成—CH—NH2面钝,—CO—OH面尖的桃心形状的空间结构。这就是极性蛋白质分子空间形变的内在力场作用根源。其它更复杂的多氨基酸结合成的蛋白质分子,会也对应的更复杂的空间结构,但形变的作用原理都是一样的。
各酸碱性氨基酸应数量与种类不同,其不同排列组合形成的蛋白质分子可以有无穷多种,于是蛋白质分子的极性形态也是多种多样的。如有强酸—弱酸型·极性蛋白质分子,有强酸—中性型·极性蛋白质分子,有强酸—弱碱型·极性蛋白质分子,有强酸—强碱型·极性蛋白质分子;又有弱酸—中性型·极性蛋白质分子,弱碱—中性型·极性蛋白质分子,弱碱—强碱型·极性蛋白质分子,等等不一而足。又如有单涡轴极性蛋白质分子,有多涡轴极性蛋白质分子,等等。这些蛋白质分子的振动力场与流动力场在分子周边空间分布不均及强弱不同,并有各自特征,导致蛋白质分子对外界环境的不同振动力场与流动力场产生相应作用,表达为特定的功能。而所有功能,都是通过振动力场与流动力场产生作用的。