高尔基体是真核细胞中的细胞器,由数个扁平囊泡堆在一起形成的高度有极性的细胞器。扁平囊为圆形,边缘膨大且具穿孔。一个细胞内的全部高尔基体,总称为高尔基器。一个高尔基体常具5–8个囊,囊内有液状内含物。高尔基体是一个囊泡系统,它由扁平膜囊、大囊泡、小囊泡三个基本成分组成,均是由光面膜构成。
扁平膜囊是高尔基体最富特征性的结构组分。在一般的动、植物细胞中,3~7个扁平膜囊重叠在一起,略呈弓形 。弓形囊泡的凸面称为形成面,或未成熟面与顺面;凹面称为分泌面,或成熟面与反面。小液泡散在于扁平膜囊周围,多集中在形成面附近。一般认为小液泡是由临近高尔基体的内质网以芽生方式形成的,起着从内质网到高尔基体运输物质的作用。
高尔基体的主要功能将在内质网上合成的蛋白质进行加工、分拣、与运输,然后分门别类地送到细胞特定的部位或分泌到细胞外,是完成细胞分泌物(如蛋白)最后加工和包装的场所。

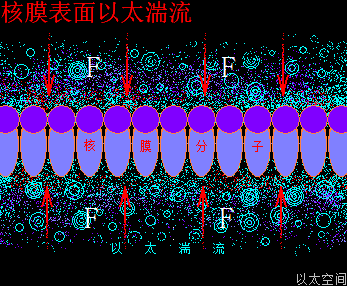
高尔基体,是真核细胞中内膜系统的组成之一。中心体位于其凹面中心。凡是膜结构,表面都存在分子以太旋涡振动发散后导致的以太湍流层,起到电荷吸附作用。
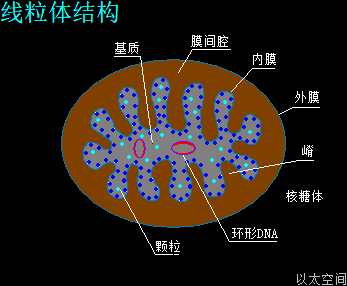
细胞的所有膜结构,其本身都不具有自主性,都是依附性存在,或者说是伴生存在。如细胞核膜必须依附在核仁周边空间,细胞膜依附在细胞核周边空间,线粒体内外膜依附在环形DNA周边空间。这一现象本身是膜中心的核心物质如核仁、细胞核、环形DNA等等的高能量水平决定的。各类膜的成分本身没有硬骨架保持膜这一曲面或展平形态的能力。
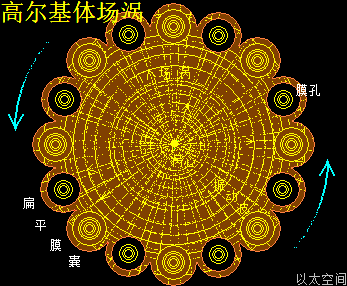
高尔基体,被有的科学家认为它是由滑面内质网进化而来的。这是认识不到生命运行机理的错判。其实高尔基体,是中心体的外围包裹物,与内质网是细胞核的外围包裹物是有一样的地位,是依附于中心体存在的。作者会在“高尔基体的形成”介绍高尔基体的诞生过程。这里只描绘高尔基体的以太波流一体形态。
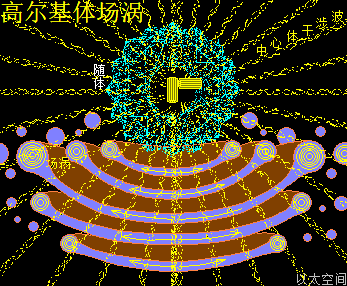
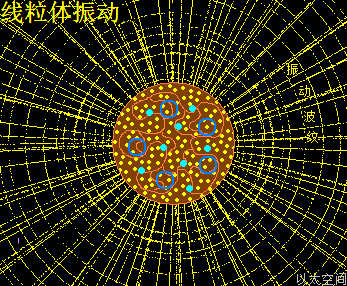
高尔基体的中心位置是中心体,中心体的主涡轴正对着穿过高尔基体的圆形中心,如雨伞伞柄穿过伞面,是为高尔基体的中心轴。中心体的振动发散与干涉波穿透高尔基体空间,在交接面会带来高尔基体膜空间的场涡运动与以太涡旋分布。此处的场涡运动范围与以太涡旋半径与中心体的干涉波束直径相当,要密集与细腻。
高等植物细胞没有中心体,却一样有高尔基体,在于植物高尔基体凹面的中心位置,即对应的动物细胞高尔基体的中心体位置处,其实也存在一种类似中心体功能,且结构也相近的蛋白质分子为主要成分的物质形态,只是受植物细胞氤氲状态的影响,其体积远小于动物细胞的中心粒与中心体,除了在光学显微镜下观察不到之外,在电子显微镜也似乎观察不到,从而被研究者忽略并未见报道于生物学知识体系中。这种物质形态,可以叫作植物中心超微粒,多个这样的超微粒一起构成植物中心超微体,在植物有丝分裂中起主导作用,并形成丝管能牵引染色体,也是高尔基体能产生一个分离的、独立完成不同功能的反面管网结构的物质作用核心。作者会在细胞分裂章节继续介绍这种植物中心超微粒与超微体。

同时,在远中心轴的扁平囊边缘位置,细胞核的振动波与干涉波,也穿透高尔基体空间,在交接面也带来高尔基体膜空间的场涡运动与以太涡旋分布。此处的场涡运动范围与以太涡旋半径与细胞核的干涉波束直径相当,要宽大与粗广。
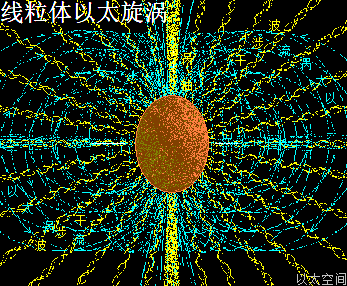
高尔基体膜内空间,主要承受中心体的振动能量,以中心轴与膜交接为中心,向膜边缘发散,并在整个膜空间形成一个大场涡,及在膜边缘形成多个小场涡。大小场涡各自对应一个大以太旋涡与多个小以太旋涡。大以太旋涡同时牵引高尔基体作绕中心轴的自转运动,小以太旋涡不断翻转后,形成小以太旋涡球,被膜包裹及被振动能量继续往外推离中心轴,最终形成小泡而脱离高尔基体。
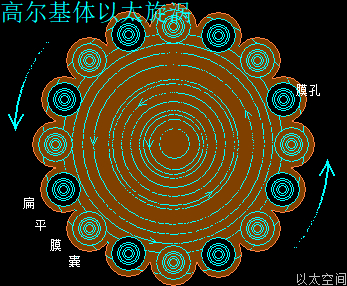
在细胞核干涉波束穿过的区域,细胞核场涡与高尔基体场涡叠加,其能量波动水平高于囊膜分子的耦合结构强度与范德华力吸引,从而破坏囊膜结构形成空洞。

高尔基体的膜场涡与膜以太涡旋,囊场涡与囊以太旋涡,及膜表面以太湍流与振动波发散,共同构成高尔基体的以太波流一体形态。