液泡是由单层膜与其内部的细胞液组成的细胞器,主要存在于植物细胞中。低等动物特别是单细胞动物的食物泡、收缩泡等也属于液泡。 液泡不是植物细胞的特有结构,只不过是植物细胞的液泡较大、液泡之间差别也较大;而动物细胞的液泡较小、液泡之间差别也不显著;或有的动物细胞的液泡不明显。
动物液泡是动物细胞内氧化还原的中心,是物质,尤其是蛋白质,如酶原粒、卵黄粒、顶体(穿孔器)等浓缩、凝结的场所。
生物学界于上世纪30年代提出了液泡系的概念,它包括高尔基液泡、溶酶体、圆球体、微体、自体吞噬泡、残质体、胞饮泡、吞噬泡、糊粉泡、中央泡、收缩泡等。认为凡是由膜包围的小泡或液泡都可算做液泡系内,它们是动植物细胞的组成分之一。同样生物学界对液泡的形成机制是一头雾水的。这里只介绍最具代表性的植物细胞的液泡的以太旋涡理论下的波流一体形态。
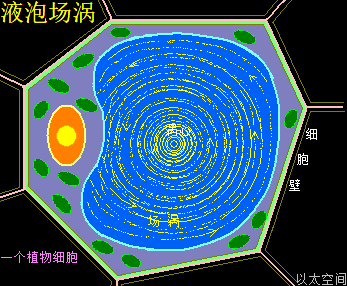
成熟的植物细胞中经常都有一个大的充满液体的中央液泡,是在细胞生长和发育过程中由小的液泡融合而成的,是单层膜包围的充满水液的泡,其中主要成分是水。植物液泡中的液体称为细胞液,其中溶有无机盐、氨基酸、糖类以及各种色素,特别是花青素等。不同种类细胞的液泡中含有不同曲物质,如无机盐、糖类、脂类、蛋白质、酶、树胶、丹宁、生物碱等,并处于高渗状态,使细胞处于吸涨饱满的状态。
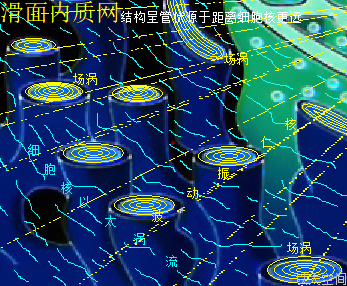
植物细胞的液泡,以下简称液泡,是存在单膜系统的泡状结构,中间有内容物。液泡膜是由蛋白质分子以太旋涡与脂类分子以太旋涡构成,形成一个封闭区域结构,其分子以太旋涡与内容物分子以太旋涡时刻处于能量振动发散状态之中。振动波在膜内外表面产生以太湍流,并在这个封闭区域不断产生波反射,导致液泡内部空间,也与肥皂泡内部空间一样,存在一个不被观察的场涡运动形态。只是这个场涡运动存在于液体之中,而不是如肥皂泡一般存在于气体中,从而有更高稳定性。场涡运动促使液泡成为一个球形结构,同时受细胞内部空间其它物质与细胞器的振动力场与流场的影响,这个球形结构产生形变,进而成为人们观察到的囊状结构。
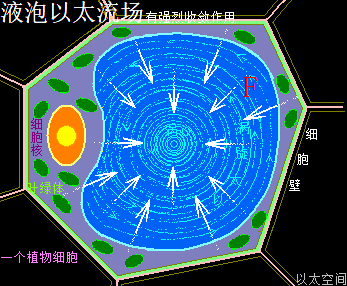
植物细胞的液泡的内容物,按现代科学研究发现其主体除了是水之外,还有无机盐、糖类、脂类、蛋白质、酶、树胶、丹宁、生物碱,等等,其实这些物质的振动力场与流动力场有统一性,即都是流动力场大于振动力场的物质运动形态,从而让液泡内部空间的整体流动力场大于振动力场,由此也导致有强烈的收敛作用。这个收敛作用,让液泡对外界表达出高渗透压作用。高渗透压作用与将一个普通容器抽成真空后表达出的压力传递类似,都是中心压强极低,从而让外界物质有趋向中心的运动状态。不是液泡外界有高压力让物质趋向液泡内部空间,而是液泡中心有超低压而让物质分子趋向液泡内部空间。这种超低压就是以太流动力场远大于振动力场后,牵引外界物质趋向中心的信号特征。
也因此,液泡内部空间的蛋白质分子,主体以碱性蛋白质分子为主,氨基酸主体也是以碱性氨基酸分子为主,其它如钾化合物离子、糖分子、生物碱,都是这么一种流场大于振动力场的物质作用形态。这些物质分子周边空间,都处于一边振动一边产生以太旋涡的波流一体状态。
液泡内容物与膜振动在液泡内部空间产生场涡与以太旋涡的同时,也让液泡处于一个低于细胞内部空间能量水平的的低以太波动频率环境之中,这就是液泡的氤氲状态。若说细胞质的氤氲状态是常温,液泡的氤氲状态有点类似冰窖低温。
液泡膜表面振动与以太湍流,内容物分子以太旋涡振动与周边以太涡流,共同构成液泡的以太波流一体。