在“细胞空间波流一体章节”中,作者粗略介绍了整个细胞空间各主要细胞器的波流一体运动形态,是各细胞器空间以太旋涡流动与以太振动波动发散形态。以太波动传递到细胞内外空间,会在辐射线程上留下场涡运动,场涡运动驱动以太形成微旋涡结构,这些微旋涡结构相互耦合,除了整体形成细胞器以太旋涡之外,还形成细胞辐射线程上的细以太涡管。如中心体细以太涡管、细胞核细以太涡管、线粒体细以太涡管,等等。这种描绘,是从各个细胞器作为完全的独立单元,来考查细胞器以太波流一体时的运动特征与结构形态。但每个细胞器,都不是完全独立单元,而是处在细胞空间这一环境里并相互临近,各自的以太旋涡、以太波动、细以太涡管分别会相互影响,因此又有不一样的运动特征与结构形态描绘。
这些细胞器的以太旋涡、以太波动、细以太涡管等,由于处在细胞整个内部空间,受细胞膜约束,存在各细胞器以太旋涡之间的相吸相斥作用、以太波动之间的相互干涉反射,细以太涡管之间的相互穿插干扰,从而出现不同细胞器之间的以太旋涡耦合作用与结构,不同细胞器之间振动波辐射的干涉波与空间干涉栏栅,及不同细以太涡管之间的相吸互联重叠作用。这些相吸相斥、相互干涉干扰、互联重叠作用,形成的不同空间以太运动形态,反过来又制约细胞器的以太旋涡流转形态、以太波动方向强弱、以太涡管粗细延伸,及细胞器在细胞内部空间的位置,等等,最终相互之间达到物质作用平衡,这就是显微镜下人们观察到细胞器在细胞空间里的稳定分布形态。
显微镜下看得到的是各细胞器在细胞空间里的分布,看不到的各细胞器之间的相互平衡作用,及以太旋涡、以太波动、细以太涡管等。这也是现代科学借助于仪器带来的窘境:仪器最高精度之下的物质结构运动,无法通过仪器信号在宏观上借光与影与人类感知,从而产生偏差及错误认知。上以下为基,宏观以微观为基,当不能正确认识微观及以下尺度物质作用形态时,宏观、宇观观察到的物质形态,都只会停留在表象层次。若不能将各表象通过自洽逻辑手段结合形成整体物质构架,就会被表象所迷惑,让科学研究陷入停滞。生命科学最重要的研究工具之一是各类光学、电子显微镜,借助显微镜,人类通过光与影认识到了微观生命及其内部结构形态,显微镜的最高精度也制约了微观生命之下的更微观物质结构形态的展现。
各细胞器应以太波动向外发散,形成空间场涡运动分布,尔后形成相应的细以太涡管。相互临近的细以太涡管之间由于相吸相斥作用,会形成空间耦合结构。不同细胞器的各自细以太涡管有长有短,管半径有大有小,涡管上的以太涡流速度有快有慢,不一而足。这不同细以太涡管之间的耦合结构,有很多种结构形态,诸如分叉结构,空间网络互联结构,大小涡管相互嵌套结构,顺逆涡管互绕结构,等等。
如分叉结构,一般是相对粗以太涡管为主杆,耦合相对细以太涡管,而相对细以太涡管再耦合相对微以太涡管,如此形成树丫状的分叉结构形态。又如相近半径、相近流转强度的细以太涡管之间,可以耦合成为网络结构,象渔网那样的结构形态。又如相同流转方向,但流转速度不同且管半径大小不一的细以太涡管,可以耦合成嵌套结构,流转速度高但管半径小的细以太涡管居中,而流转速度慢但管半径大的细以太涡管包裹其外,如现实中大水管套小水管那般结构形态。又如相互临近但互为顺逆的两条细以太涡管,会产生互绕耦合形态,如两条藤蔓互绕延伸,等等。这些树丫状、网络状、嵌套状、互绕状的细以太涡管耦合结构,又相互联结耦合,形成复杂的细胞空间以太涡管立体分布形态。
细胞整体空间里的各细以太涡管形成的涡管耦合结构,会有流体涡管的一般特征,有如龙卷风一般的结构形态与空间物质流转模式,如涡管壁上的以太流速最高,并随半径增加而流速不断减小,表达为以太涡管流场,这是细胞器整体以太旋涡流场之下的另一种局部流场形式。又如涡管中心的以太流速度最低,甚至处于相对静止状态;以太涡管两端分别存在以太涡口,一头在细胞器内部空间,一头一细胞器周边空间,涡口能吸引其它物质,涡管则成为其它物质的运输通道,等等。以太涡管还与以太旋涡一样,应流转方向有顺逆之分,从而不同涡管之间能产生相吸或相斥作用。这些以太涡管互联耦合网状结构,还应细胞器的活性、生命周期,也存在产生、成长、稳定、败坏、消失等等过程。
各细胞器形成的细以太涡管中心,由于涡管壁的流速远大于涡管中心,形成一个封闭时空,当细胞器中心的振动波发散时,有部分振动波会在细以太涡管内壁反射并沿涡管通道传递,如可见光在玻璃光纤内壁反射与通道传递一般,表达为振动波高效通过细以太涡管内部空间发散到细胞器周边空间,同时细胞器周边空间的其它振动能量波,也可以高效地通过细以太涡管传递到细胞器中心,从而影响细胞器周边空间物质分布与细胞器内部空间的结构与细胞器生理活动。
细胞整体空间里,各细胞器的高效传递振动能量波的以太涡管互联互通耦合网络结构,就是细胞气脉。
细胞空间过于微小,而以太波流一体形态又过于精微,只能通过感悟与想象来理解。要直观认识这种细胞气脉分布形态,可以参考现实中一种叫等离子球的玩具,又叫静电球、闪电球,即通上电,会产生电弧分叉四处逸散现象。这等离子球内部空间的电弧分布形态,与细胞器以太涡管互联耦合网络分布形态,即细胞气脉,有几近一样的运动特征与物质结构形态,在于两者的产生机理几近一致。其它如借用树叉分形形态、叶脉网状分布形态、闪电分叉传导形态、河流分叉网状分布形态、神经末梢分叉分布形态,等等,都可以直观理解细胞器周边空间以太涡管互联耦合网状分布结构。这也是宇宙全息的体现,无论是微观、宏观、宇观,都有相近相同的物质运动特征与物质结构形态,不同时空尺度可以相互印证理解。
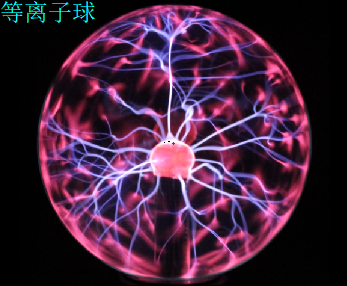
细胞气脉,是各细胞器以太涡管耦合互联结构,能高效传递各细胞器的振动能量,让各细胞器之间实现互联互通,并实现高效物质运输、快速能量互换,是各细胞器之间的高速公路。就如现实中各城市乡镇之间的交通运输网络道路。细胞气脉,是现实科学没有认识到最重要的细胞结构形态之一。
在细胞空间里的细胞器层面上,最高能量波动强度的细胞器是中心体、细胞核、线粒体(对植物来说,是叶绿体、细胞核、线粒体),如此细胞气脉对应的是中心体细以太涡管、线粒体细以太涡管、细胞核细以太涡管这三种细胞器细以太涡管为主导的互联互通耦合结构。这三种细胞器各自的细以太涡管优先发生耦合作用,形成耦合互联结构,再相互耦合互联,分布在整个细胞内部空间,成为细胞气脉主体,然后耦合其它细胞器细以太涡管,如溶酶体以太涡管、液泡以太涡管等,填充整个细胞内部空间,甚至发散到细胞外部空间,等等,是为细胞气脉整体。作者就中心体、细胞核、线粒体这三种细胞器各自的细以太涡管耦合互联结构分别作一个独立小节介绍。
中心体、线粒体、细胞核,各有一个以太旋涡,每个以太旋涡两极都有一条大以太涡管及一个以太涡口,如此这些大以太涡管之间,优先发生耦合联结作用,其相互联结形成的以太涡管通道,就是细胞气脉的主杆,高效传递中心体振动波、线粒体振动波、细胞核振动波,让中心体中心、线粒体中心、细胞核中心之间实现能量交换,并相互影响自的生化反应,会在“细胞生长与分裂”作一个解析这种能量交换形态。