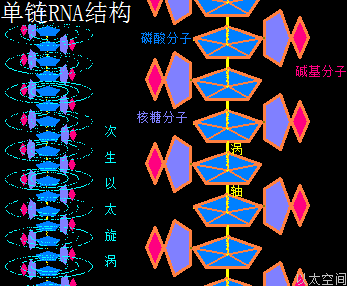
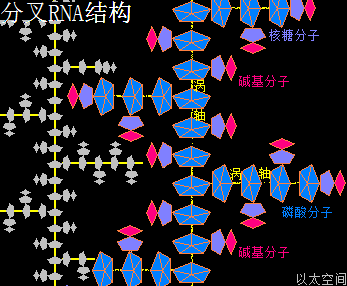
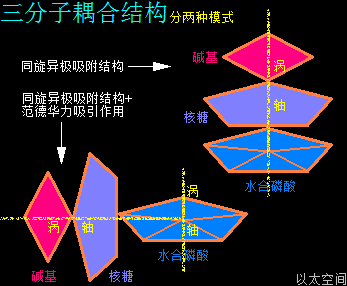
核糖核酸RNA与脱氧核糖核酸DNA,其组成单元分别是核糖核苷酸分子与脱氧核糖核苷酸分子,两种分子各有四种碱基组合形态,共五种碱基类型。所带的不同碱基分别是核糖核苷酸分子由四种碱基,即腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)构成的,脱氧核糖核苷酸分子则由四种碱基,即腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、胸腺嘧啶(T)构成的。这里分别就这五种碱基作一个以太旋涡理论下的模型结构介绍。
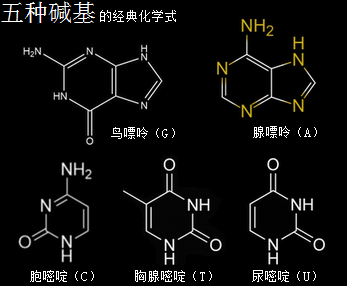
——>腺嘌呤(A)
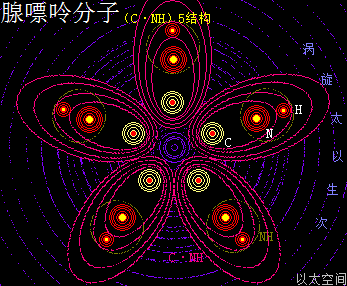
腺嘌呤,就是维生素B4,又称6-氨基嘌呤,化学分子式为C5H5N5,碱基代码为A。
以太旋涡理论下,腺嘌呤的化学结构式为(C·NH)5,即先五个N与五个H耦合成为五个—(NH),再五个C与五个(NH)耦合成五个(C·NH),最后五个(C·NH)耦合成为(C·NH)5。
在腺嘌呤分子以太旋涡中,五个(C·NH)以在同一平面上通过范德华力作用耦合在一起及互绕,(C·NH)中的C与N通过异旋同极吸附作用耦合在一起,极限偏向状态就如一个五角星形态,并在周边空间形成次生以太旋涡,及相应的流场与振动力场。
可以看出腺嘌呤分子以太旋涡与核糖分子以太旋涡,有几近一样的空间结构形态与运动模式。又由于核糖分子中的碳原子比腺嘌呤分子中的碳原子多一个氢原子,导致核糖分子以太旋涡半径与流场范围比腺嘌呤分子以太旋涡半径与流场范围要来得大一些,但分子结构的稳定性则要差一些。
NH就是亚氨基离子以太旋涡,比—NH2少一个氢原子,其振动力场大于流动力场,其与C原子耦合后形成(C·NH)离子以太旋涡,其振动力场及流场与—NH2—CH的振动力场及流场有相近形态。(C·NH)就是一个弱化形态的—NH2—CH。由此带来的嘌呤碱基分子有氨基的作用特征,但作用强度相对氨基要弱。又由于在一个嘌呤碱基分子上存在五个(C·NH),因此作用频率要相对氨基要高。
——>鸟嘌呤(G)
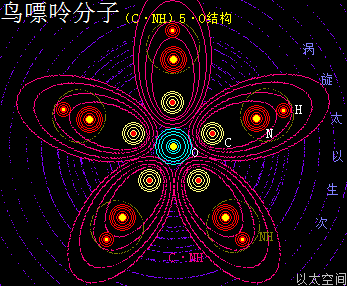
鸟嘌呤的化学分子式为C5H5N5O,碱基代码为G。
以太旋涡理论下,鸟嘌呤的化学结构式为(C·NH)5·O,即先五个N与五个H耦合成为五个(NH),再五个C与五个(NH)耦合成五个(C·NH),及后五个(C·NH)耦合成(C·NH)5,最后(C·NH)5与一个O耦合成(C·NH)5·O。
在鸟嘌呤分子以太旋涡中,五个(C·NH)以O为核心在同一平面上通过范德华力作用耦合在一起及互绕,极限偏向状态就如一个五角星形态,并在周边空间形成次生以太旋涡,及相应的流场与振动力场。
可以看出鸟嘌呤比腺嘌呤在次生以太旋涡涡心处多一个O原子以太旋涡,其它结构均一致,因此其振动力场与流动力场形态都要强于腺嘌呤。
——>胞嘧啶(C)
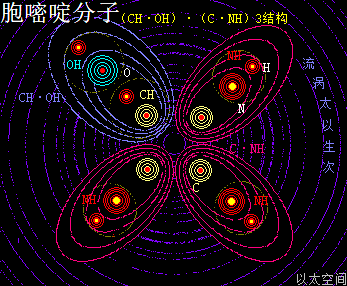
胞嘧啶,学名为4-氨基-2-羰基嘧啶,化学分子式为C4H5N3O,碱基代码为C。
以太旋涡理论下,胞嘧啶的化学结构式为(CH·OH)·(C·NH)3,即先三个N与三个H耦合成三个(NH),一个C与一个H耦合成为一个(CH),一个O与一个H耦合成为一个(OH);再三个C与三个(NH)耦合成为三个(C·NH),及一个(CH)与一个(OH)耦合成为一个(CH·OH);最后三个(C·NH)与一个(CH·OH)耦合成为(CH·OH)·(C·NH)3。
在胞嘧啶分子以太旋涡中,一个(CH·OH)与三个(C·NH),在同一平面上通过范德华力作用耦合在一起及互绕,其中(CH·OH)中的O与C通过异旋同极吸附作用耦合在一起,极限偏向状态就如一个十字形态,并在周边空间形成次生以太旋涡,及相应的振动力场与流动力场。
就如NH是—NH2的弱化版,(CH·OH)也是—CO—OH,即羧基的弱化版,其流动力场大于振动力场,由此带来的嘧啶碱基分子有羧基的作用特征,但相对羧基要弱。
——>胸腺嘧啶
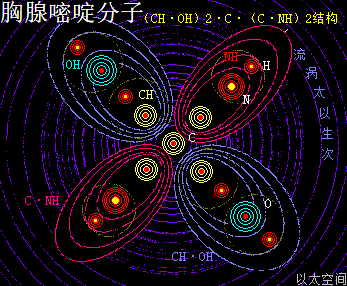
胸腺嘧啶,化学分子式为C5H6N2O2,碱基代码为T。
以太旋涡理论下,其胸腺嘧啶的化学结构式为(CH·OH)2·C·(C·NH)2,即先二个O与二个H耦合成为二个(OH),二个N与二个H耦合成为二个(NH),及二个C与二个H耦合成为二个(CH);再二个(CH)与二个(OH)耦合成为二个(CH·OH),及二个C与二个(NH)耦合成为二个(C·NH);再后二个(CH·OH)与二个(C·NH)及最后是一个C,耦合成(CH·OH)2·C·(C·NH)2。
在胸腺嘧啶分子以太旋涡中,四个离子以太旋涡,即两个(CH·OH)与两个(C·NH),在同一平面上通过范德华力作用耦合在一起,以一个C原子以太旋涡为平面中心互绕,极限偏向状态就也如一个十字形态,并在周边空间形成次生以太旋涡,及相应的流场与振动力场。
相对胞嘧啶,胸腺嘧啶少了一个(C·NH),多了一个(CH·OH)及C原子,因此胸腺嘧啶的以太流场比胞嘧啶的以太流场要更强,但胸腺嘧啶的振动力场比胞嘧啶的振动力场要弱。这一强流场形态让胸腺嘧啶有更高吸引与收敛作用。
——>尿嘧啶(U)
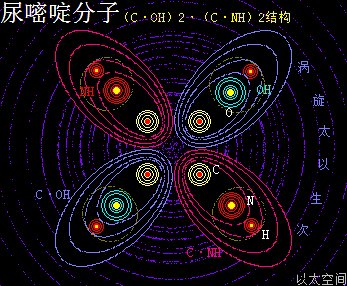
尿嘧啶 ,化学分子式为C4H4N2O2,碱基代码为U,是RNA特有的碱基。
以太旋涡理论下,尿嘧啶的化学结构式为(C·OH)2·(C·NH)2,即先二个O与二个H耦合成为二个(OH),及二个N与二个H耦合成为二个(NH);再二个C与二个OH耦合成为二个(C·OH),及二个C与二个(NH)耦合成二个(C·NH);最后是二个(C·OH)与二个(C·NH)耦合成(CH·OH)2·(C·NH)2。
在尿嘧啶分子以太旋涡中,四个离子以太旋涡,即两个(C·OH)与两个(C·NH),在同一平面上通过范德华力作用耦合在一起及互绕,极限偏向状态就也如一个十字形态,并在周边空间形成次生以太旋涡,及相应的振动力场与流动力场。
(C·OH)也是—CO—OH,即羧基的弱化版,其流动力场大于振动力场,由此带来的尿嘧啶有羧基的作用特征,但相对羧基要更弱。
(CH·OH)2·C·(C·NH)2即胸腺嘧啶,可以看出尿嘧啶比胸腺嘧啶在次生以太旋涡涡心处少一个C原子以太旋涡,并在两个(CH·OH)结构中各少一个H原子,其它结构均一致,于是相比之下胸腺嘧啶有相对较高强度的振动力场与流动力场形态,但结构稳定性相对要差。反之尿嘧啶的结构稳定性要高些,可以在更高的环境振动力场中保持原有形态。这也是胸腺嘧啶可以转换为尿嘧啶的物质作用根源,会在后续基因原理章节继续论述两种嘧啶的转换机制。
(CH·OH)·(C·NH)3即胞嘧啶,可以看出尿嘧啶比胞嘧啶少一个(C·NH)与一个(CH·OH),多两个(C·OH),而(C·OH)是(CH·OH)的弱化结构,因此尿嘧啶流场相对胞嘧啶流场要强,但尿嘧啶振动力场相对胞嘧啶振动力场要弱。
由以上五种碱基的以太旋涡理论下的全新构建与描绘,可以为正确认识与理解基因运作机制提供理论支持。这五种碱基的振动力场与流场各自对比呈一个梯度分布:
振动力场梯度分布
鸟嘌呤(G)>腺嘌呤(A)>胞嘧啶(C)>胸腺嘧啶(T)>尿嘧啶(U)
流动力场梯度分布
胸腺嘧啶(T)>尿嘧啶(U)>胞嘧啶(C)>鸟嘌呤(G)>腺嘌呤(A)