四季,即春夏秋冬四个季节,一般感官是春季温润,夏季炎热,秋季凉爽,冬季寒冷,被认为是地球在围绕太阳公转时,由于黄道面与赤道面存在相交角度,造成太阳直射点在地球南北纬23°26′之间往返移动形成以年为周期的往复变化,从而引起正午太阳高度变化和昼夜长短变化,进而影响温度、气候,等等,是为四季更替。每三个月为一季。中国传统上是以“二十四节气”立春、立夏、立秋、立冬分别作为春季、夏季、秋季、冬季的起始。
虽然黄道面与赤道面存在相交角度结合地球绕太阳公转形成四季极直观,作为四季成因也是大差不差的,但科学界由于缺失对以太存在的认知,导致对四季背后的物质作用机制并没有一个根源上的诠释,这种只用地轴偏向与阳光多寡来作为四季成因,是流于表面。上面几个小节已经用天气、地气、空气、水气等概念来重新解构地表空间气温气候的影响。这里更详细说明这些不同形态的以太波流一体的影响过程,与四季背后更深层次的物质作用机制。
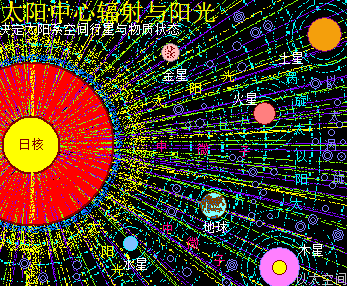
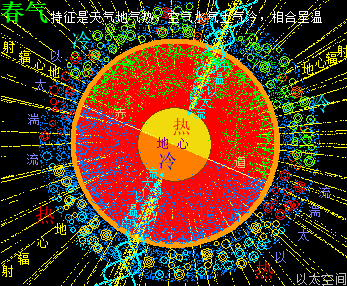
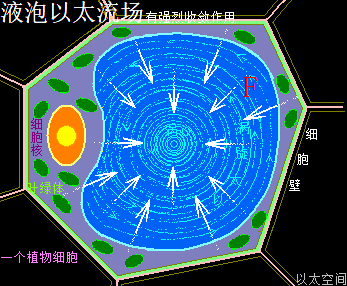
太阳系是宇观恒星级别的以太旋涡,其中心日核随时在进行超巨量核聚变活动,其中心辐射能量表达为中微子这种超高频波辐射,以球面膨胀形式向太阳系空间扩散,同时太阳光球层的原子以太旋涡激烈振动产生可见光、红外线、紫外线,也向太阳系空间扩散。由于辐射能量是呈立体空间分布的关系,存在以距离三次方的衰减梯度分布。如此太阳中心辐射与太阳光,共同构成太阳辐射,导致太阳系空间的以太随之波动,产生超巨量微以太涡旋。整个太阳系空间,就是太阳辐射能量波动的空间,也是微以太涡旋流动的空间,也是以太之海,这是太阳系的以太波流一体,全方位影响太阳系空间所有行星与原子的活跃性。
离太阳中心有不同的距离,有不同的辐射作用强度与不同的微以太旋涡分布密度。对地球来说,通过接受太阳中心日核辐射,从而强化地心地核能量波动;通过接受太阳光,从而强化地球表面空气能量波动;通过南北两极以太涡口吸收公转轨道上以太微涡旋落入地心,从而让地球维持核聚变,并不断成长并产生地心能量辐射。其中太阳中心的日核辐射是全方位的全空间作用的,太阳光辐射只影响能被太阳光照射到的地方,照射不到的地方就不能产生影响。以太涡口对以太微涡旋的吸收则局限于两极上方空间。
这是太阳系波流一体对地球表面四季的影响。其太阳中心能量辐射对地表的影响是最根源所在。无论是太阳光还是微以太涡旋,及地心能量辐射,都是太阳中心能量辐射后带来的系列影响与连锁传递。
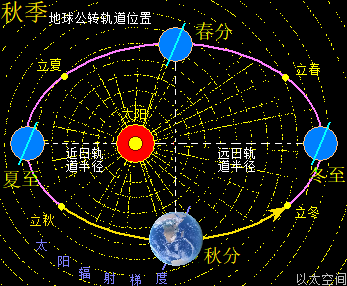
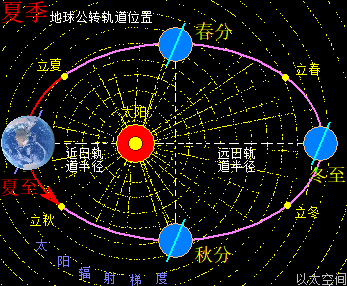
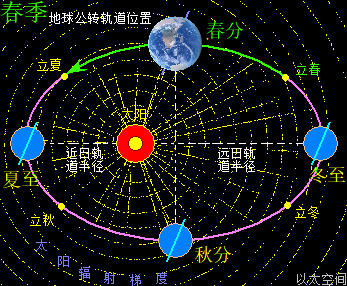
地球与月球周边空间都存在一个宇观行星级别的以太旋涡,地球受月球引力影响导致地球以太旋涡产生偏向流转形态,并构成地月系。地月系是宇观层次的极性双宇观行星以太旋涡的次生以太旋涡耦合结构。这以太旋涡是一个偏向性的涡流循环运动形态,即涡流以椭圆形态而非正圆形态流转。地球公转其实是地球以太旋涡在太阳以太旋涡的涡流上作漂移运动,结合地球以太旋涡偏向,从而在公转过程中也随之出现椭圆形公转轨道形态。
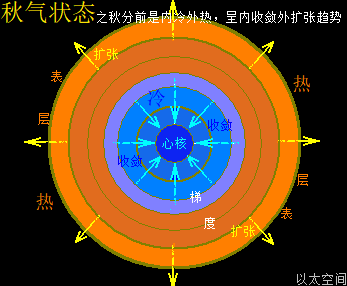
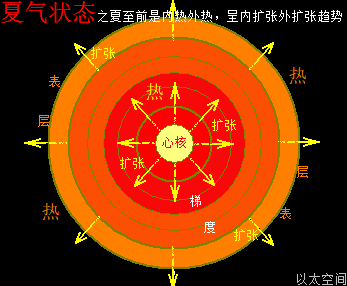
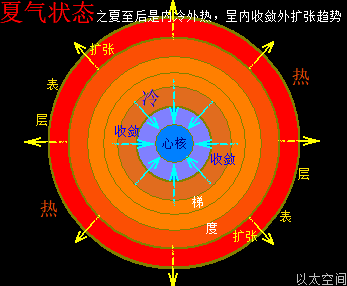
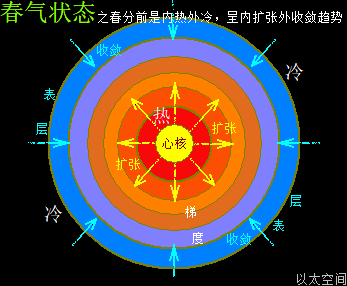
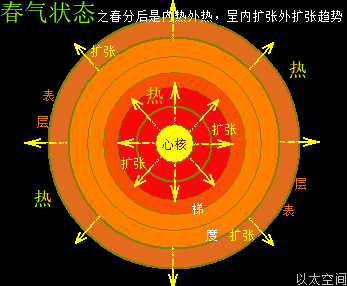
地球这种椭圆形公转轨道,让地球公转时,在一年周期内,出现轨道半径靠近太阳与远离太阳两种运行状态。当轨道半径缩短靠近太阳时,地心接受到的太阳辐射能量会增强,从而强化地心能量波动,继而强化地气强度,并让空气、水气、土气的能量波动强度提升。反之,当轨道变长远离太阳时,地心接受到的太阳辐射能量会减弱,从而削弱地心能量波动,继而弱化地气强度,并让空气、水气、土气的能量波动强度下降。这才是四季的真正的主要内在物质作用机制。作者会后面“春气”、“夏气”、“秋气”、“冬气”四个小节专门作详细解析。
应月球绕地球公转,月球位置会处于太阳与地球之间,及太阳与地球之外的以月为周期的分布情况。如此地月系的重心,并不是地球地心,而是位于地球与月球之间的连线上靠近地球的某个点上。地月系重心不在地球地心上的结果,是地球一边自转并受月球公转,一边绕太阳公转时,地心的运动轨道不再是一个椭圆形轨迹,而是以月为周期的轨道半径最远离太阳(阴历初一),及轨道半径最靠近太阳(阴历十五)的,以重心椭圆形轨迹为核心线的波浪起伏—椭圆形轨迹。地月系绕太阳公转,如一个在空中飞转的两头不一样大小的哑铃。这也导致地球表面出现正午远离太阳与靠近太阳的以月为单位的周期性变化。这种正午远离太阳与靠近太阳的周期性变化,带来的结果是地表接受的阳光强度会以月为周期的增加—减小—增加的往复循环。这种阳光强度的细微变化难以度量,但可以让人的身体到能感受到略微差异,比如每逢初一日子会感觉温度略显冷清,而在十五日子会感觉温度略显温暖,进而影响心情,自然,这种感觉是应人而异,有的人感受敏锐,有的人感受迟钝,带来的影响是不一样的,也是应具体的天气、地理等不同,并不是绝对感觉到暖或冷。当处于夏季或冬季时,这种月球影响下的正午高度变化带来的地表接受的阳光的强度会被放大,于是这种感觉就会更显著。如此,四个季节都存在以月为周期的接受阳光变大变小的过程,影响生命活动与细胞复制。

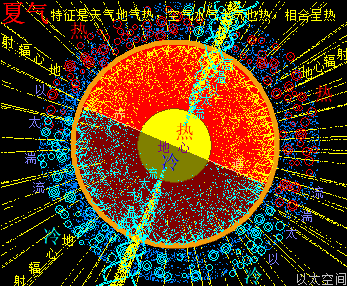
而地轴偏向,在夏冬两季会导致地球南北两极的以太涡口,分别处于完全阳光之中,与完全阴暗之中,对应的是南北极地分别出现极昼与极夜现象。当北半球处于极昼状态时,北极以太涡口吸入的微以太涡旋能量是一种高频能量波动,同进南半球处于极夜状态,南极以太涡口吸入的微以太涡旋能量是一种低频能量波动。
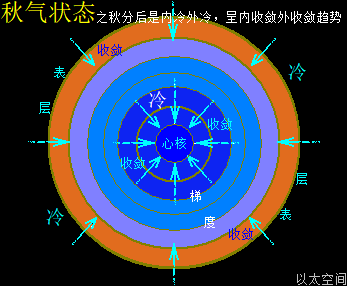
北极以太涡口吸入高频能量微以太涡旋时,北半球的半个地心的活跃性增加,核聚变释放的能量强度也随之增加,从而导致北半球的地幔,以涡轴为中心线的四周温度升高,地幔熔浆流速增加,并产生强红外辐射,及北半球地表地气强度增加,最终导致北半球地表空气、水气、土气的能量波动水平都跟着增加,宏观上表达为升温,与春季、夏季的到来。同时,南极以太涡口吸入低频能量微以太涡旋时,南半球的半个地心的活跃性减弱,核聚变释放的能量强度也随之减小,从而导致南半球的地幔,以涡轴为中心线的四周温度降低,地幔熔浆流速减小,并只产生弱红外辐射,及南半球地表地气强度减小,最终导致南半球地表空气、水气、土气的能量波动水平都跟着减小,宏观上表达为降温,与秋季、冬季的到来。
南北半球地心与地表的这种能量增加与减小动态过程,是以年为周期的相互轮换过种,其中以季为四分之一周期,通过温度与人的体感来划分出春夏秋冬四个季节。两个半球的地心活跃性此消彼涨的过程,让两个半球还出现体积上一扩张一收缩的变化,犹如人的心脏分左右心房心室的跳动过程,这是一个以年为单位的宇观行星级别的心脏的跳动频率。地心跳动与人心跳动,有完全对应的诸多如心率、振动波、体液流动等等属性的描绘。这仍是宇宙全息的体现。
南北两极以太涡口吸入低高频能量的以太涡旋,除了通过地球两极以太涡管深入地心,影响地心核聚变外,还直接影响两极极地寒流的强度。当北半球处于极昼状态时,极地寒流的强度受高频能量以太的作用,寒流的温度要相对高,极地处于夏季,地表温度受阳光辐射也要高,对应北半球接受到的阳光整体能量高,空气温度高,于是能极大地平衡寒流的低温影响,让整个北半球的夏天的大气温度变得高。同时,南半球处于极夜状态,极地寒流的强度受低频能量以太的作用,寒流的温度要相对要低很多,极地处于冬季,地表温度没有阳光辐射而变得更低,对应南半球接受到的阳光整体能量水平低,空气温度低,于是不能有效平衡寒流的低温影响,让整个南半球的冬天的大气温度变得很低。
以上是地球的地轴偏向,导致的表面与轨道的影响。不仅仅是在公转轨道上产生正午太阳高度的变化带来的阳光强度不同,还产生两极涡口微以太涡旋能量水平有高低,及寒流强度、地心核聚变与地心辐射强度高低的差异,从而决定地表大气的温度与四季的划分。相比较于阳光,地气的强度大小变化对四季的气候变换更为重要。只要地气强度足够高,就可以让地球表面的空气、水气、土气处于高能量高温度水平。哪怕远离太阳,阳光照射强度极弱的木星、土星、海王星、天王星,都应自身地气的强度发散,可以有适宜的地表温度而让生命得以生存与繁衍,而非科学界想当然地以为这些远日行星由于缺少阳光辐射,必会是天寒地冻,毫无生机的。而过大质量的行星产生过高强度的地气辐射,其实就是恒星了。太阳中心辐射在太阳表面的分布,其实就是太阳地气,不止带来高温的太阳表面环境,还让整个太阳系都接受能量辐射而充满生机。

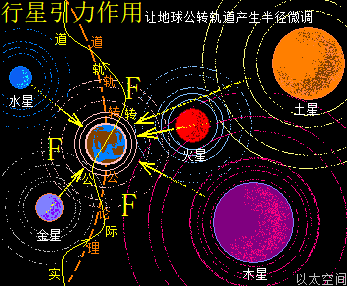
太阳系还包含九大行星,影响最大的是离地球最近的五个行星,是为水星、金星,称为地内行星,及火星、木星、土星,称为地外行星。五个行星周边空间也都各自存在一个宇观行星级别的以太旋涡。众行星绕太阳公转时,其以太旋涡的力场表达为引力,会与地球引力产生排斥作用。特别是连珠期间,如五星连珠、七星连珠等天象,众行星间的引力在一条直线上,会产生叠加作用效果,从而对地球的排斥作用特别明显。比如当水星、金星、地球位置在太阳同侧并构成一条直线时,地内行星以太旋涡间的排斥力会让地球远离太阳,于是地球此时的轨道半径会变长。当火星、木星、土星、地球位置在太阳同侧构成一条直线,地外行星以太旋涡间的排斥力会让地球靠近太阳,于是地球此时的轨道半径会变短。如此不同行星的轨道周期性变化,会让地球在五个行星的周期的最小公倍数,即六十年内,会周期性地出现地球过于远离太阳与过于靠近太阳的轨道变化。这也六十甲子纪年方式的由来,作者不展开细说。
这五大行星的引力排斥作用是随时随地发生,导致地球的轨道运行,会被随时随地地推向太阳或推离太阳,整体是一条如噪音态、不规则锯齿状的椭圆运行轨迹线。并导致地球气候的影响。
当地球远离太阳时,轨道半径变长,地表接收到的整体阳光强度会减弱,地表空气的平均温度就会下降些。同时地心受到太阳中心日核辐射能量的波动影响也会减少,表现活性不高,一如人在低温下时心脏跳动更缓。地球两极吸收到的微以太涡旋的能量水平也会降低,这些因素都导致地心的核聚变反应也会跟着变弱,于是地心辐射也会减弱,进而导致地幔红外辐射也减弱,最终也导致地表的地气强度减弱与空气温度降低。当这种轨道半径变长发生在具体的季节上,在体感上就是这个季节较往年温度要低,有冬天过于冷,夏天很凉爽的感觉。在天象上则是能看到金星、水星凌日。
金星、水星凌日又分三种情况,分别是水星凌日,金星凌日,金星、水星同时凌日。由于金星质量比水星大,且距离地球比水星近,因此当金星凌日时,地球会被金星以太旋涡的斥力推离到远太阳公转轨道,于是地球整体气候温度偏低,体感为冬天很冷,夏天偏凉。当水星凌日时,地球会被水星以太旋涡的斥力推离到略远太阳公转轨道,于是地球整体气候温度略偏低,体感为冬天偏冷,夏天偏温。当金星、水星同时凌日,地球会被两行星以太旋涡的斥力推离到很远太阳公转轨道,于是地球整体气候温度偏很低,体感为冬天极冷寒,是为寒冬,而夏天则很凉,是为凉夏。
当地球靠近太阳时,轨道半径变短,地表接收到的整体阳光强度会增强,地表空气的平均温度就会升高些。同时地心受到太阳中心日核辐射能量的波动影响也会增加,表现更有活性,一如人在高温下时心脏跳动更快。地球两极吸收到的微以太涡旋的能量水平也会升高,这些因素都导致地心的核聚变反应也会跟着变强,于是地心辐射也会增强,进而导致地幔红外辐射也增强,最终也导致地表的地气强度增强与空气温度升高。当这种轨道半径变短发生在具体的季节上,在体感上就是这个季节较往年温度要高,有冬天很暖和,夏天非常热的感觉。在天象上则是能看到火星、木星凌月。
火星、木星凌月也分三种情况,分别是火星凌月,木星凌月,火星、木星同时凌月。由于火星质量与地球相当,且距离地球比木星近,因此当火星凌月时,地球会被火星以太旋涡的斥力推离到近太阳公转轨道,于是地球整体气候温度偏高,体感为夏天很热,冬天偏暖。当木星凌月时,地球会被木星以太旋涡的斥力推离到略近太阳公转轨道,于是地球整体气候温度略偏高,体感为夏天偏热,冬天偏凉。当火星、木星同时凌月,地球会被两行星以太旋涡的斥力推离到很近太阳公转轨道,于是地球整体气候温度偏很高,体感为夏天极炎热,是为炎夏,而冬天则很暖和,是为暖冬。
土星在五星中距离地球最远,相对影响最小。单独土星凌月,对地球的影响是平和,可以作为其余四星凌日、凌月的参考基准。
五星对地球的干扰了,除了上述单星凌日、凌月,或双星凌日、凌月特殊之外,总体是相互混杂作用的。如金星凌日的同时,火星也在凌月,或金星、水星凌日的同时,火星、木星也在凌月,于是对气候的影响是这几颗行星的合力对地球公转轨道的作用结果,要具体年份、月份、日期来具体分析。
如此,整个地球公转轨道,是地球月球耦合结构的地球以太旋涡偏向后产生椭圆形轨迹,叠加上月球绕地球公转产生的以月为周期的波浪起伏状轨迹,再叠加五大行星引力作用产生的随时随地位置变化导致的轨道微调,构成地球公转轨道的实际运行形态。由此带来的轨道半径在不同日期,不同月份,不同年份,有不一样的正午太阳高度差异,影响太阳中心辐射、太阳光、两极吸引微以太涡旋的能量强度在地球空间的不同,进而促使地心核聚变烈度、寒流强度,与地表大气温度的一系列连锁反应的不同,产生每一个季节的在不同年份下的相同日期、月份,都有不同的温度、气候等等变化,最终作用于地球表面生物圈各种生命活动与细胞复制。这需要具体宇空行星分布、月球位置、不同经纬度,甚至太阳活动年,等等因素来考查季节形态。
这是地球表面一年四季现象背后的更详细的物质作用机制。
最后作一点说明:地球与水星一样,也存在进动作用,从而导致公转轨迹线并不是一个闭合曲线,而是一条螺旋收敛、结合上面几大因素叠加影响的波浪、锯齿轨迹形态。只是这个进动影响在以季为周期的季节变化上影响微乎其微,这里作者就忽略描绘,并以闭合曲线来描绘这公转轨迹。





















































.png)